Fatemeh Ghorbani1,
Elham Gheisari3,
Farhad Safarpoor Dehkordi3 ![]()
For correspondence:- Farhad Dehkordi Email: Elhamgheysari7@yahoo.com Tel:+989177142875
Received: 25 November 2016 Accepted: 8 July 2016 Published: 30 August 2016
Citation: Ghorbani F, Gheisari E, Dehkordi FS. Genotyping of vacA alleles of Helicobacter pylori strains recovered from some Iranian food items. Trop J Pharm Res 2016; 15(8):1631-1636 doi: 10.4314/tjpr.v15i8.5
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To study the vacA genotype status of H. pylori isolated from some Iranian food items.
Methods: Three hundred assorted samples of fish, ham, chicken, vegetable and meat sandwiches, and minced meat were purchased and tested using culture method. Those that were H. pylori-positive were analyzed for presence of vacA genotypes using polymerase chain reaction (PCR).
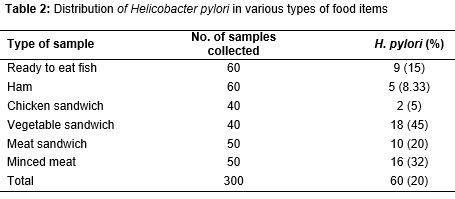
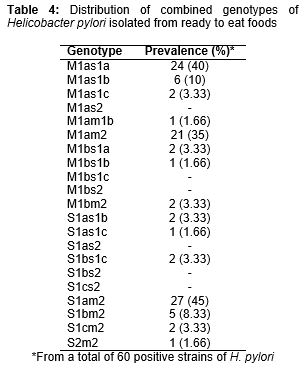
Results: Sixty out of 300 (20 %) food samples were positive for H. pylori. Vegetable sandwich (45 %), minced meat (32 %) and meat sandwich (20 %) were the most commonly contaminated. The most commonly detected genotypes in the meat-based foods, viz, vegetable sandwich and ready to eat fish, were vacA s1a, vacA m1a and vacA m2, respectively. The most commonly detected combined genotypes were s1am2 (45 %), s1am1a (40 %) and m1am2 (35 %).
Conclusion: The presence of similar genotypes in H. pylori strains of foods and those of human clinical samples suggest that contaminated foods may be the source of bacteria for humans.
Introduction
Helicobacter pylori is a microaerophilic gram-negative bacteria which is known as the causative agent of gastric adenocarcinoma, type B gastritis, peptic ulcer disease, and mucosa-associated lymphoid tissue lymphoma [1]. Recorded data show that 17 to 86 % of patients with peptic ulcers were positive for H. pylori [1-3]. Despite the high incidence of H. pylori, its exact routes of transmission and origin have not been well determined.
The role of foods in the transmission of H. pylori is still unknown but there are several studies on its isolation from various types of food like milk, vegetables and salad [4-7]. Favorable conditions for the growth of bacteria in various types of foods including ready to eat foods, meat, salads and vegetables would enhance H. pylori the survival [5,7].
To appraise the pathogenicity of H. pylori, evaluation of the status of genotypes is requisite. The most commonly known virulence markers among H. pylori strains of different clinical samples of animals and human beings are the vacuolating cytotoxin (vacA) [7-9]. The vacA gene is associated with injury to epithelial cells of gastric tract. This gene is polymorphic, comprising variable signal regions and mid-regions. The s-region is classified into s1 and s2 types and the m-region into m1 and m2 types. The s1 type is further subtyped into s1a, s1b and s1c subtypes, and the m1 into m1a and m1b subtypes [10,11]. Genotyping using vacA virulence marker is considered as one of the most effective approaches for the study of correlations between H. pylori isolates from different samples.
There were scarce published data on the genotyping of the vacA gene of the H. pylori isolated from food items. Also data about the epidemiology and routes of transmission of this bacterium are scarce too. With the high prevalence of H. pylori among Iranians [4,5,7,9] and based on the good features of foods in the survival of H. pylori [12], the present study was carried out to ascertain the status of vacA genotypes of H. pylori isolated from various types of food items.
Methods
Sample collection
From March 2013 to July 2014, overall 300 samples of food items samples including ready to eat fish (n = 60), ham (n = 60), chicken sandwich (n = 40), vegetable sandwich (n = 40), meat sandwich (n = 50) and minced meat (n = 50) were purchased from the supermarkets of Shiraz province, Iran. Samples (100 mL, in sterile glass containers) were transported to the laboratory at ca. 4 oC within a maximum of 6 h after sampling. All samples were kept under refrigeration in plastic bags; information about dates of production and of assigned shelf lives was not presented.
Isolation of Helicobacter pylori
Twenty five milliliters of each homogenized sample were added to 225 mL of Wilkins Chalgren anaerobe broth (Oxoid, UK) supplemented with 5 % of horse serum (Sigma, St. Louis, MO, USA) and colistin methanesulfonate (30 mg/L), cycloheximide (100 mg/L), nalidixic acid (30 mg/L), trimethoprim (30 mg/L), and vancomycin (10 mg/L) (Sigma, St. Louis, MO, USA) and colistin methanesulfonate (30 mg/L), cycloheximide (100 mg/L), nalidixic acid (30 mg/L), trimethoprim (30 mg/L), and vancomycin (10 mg/L) (Sigma, St. Louis, MO, USA) and incubated for 7 days at 37 °C with shaking under microaerophilic conditions. Then, 0.1 mL of the enrichment selective broth was plated onto Wilkins Chalgren anaerobe agar (Oxoid, UK) supplemented with 5 % of defibrinated horse blood and 30 mg/L colistin methanesulfonate, 100 mg/L cycloheximide, 30 mg/L nalidixic acid, 30 mg/L trimethoprim, and 10 mg/L vancomycin (Sigma, St. Louis, MO, USA) and incubated for 7 days at 37 °C under microaerophilic conditions. For comparison, a reference strain of H. pylori (ATCC 43504) was employed.
DNA extraction and Helicobacter pylori 16S rRNA gene amplification
Suspected colonies were identified as H. pylori based on the PCR technique. Genomic DNA was extracted from the colonies with typical characters of H. pylori using a DNA extraction kit for cells and tissues (Roche Applied Science, Germany, 11814770001) according to the manufacturer’s instructions and its density was assessed by optic densitometry. Extracted DNA was amplified for the 16S rRNA gene (primers: HP-F: 5'-CTGGAGAGACTAAGCCCTCC-3' and HP-R: 5'-ATTACTGACGCTGATTGTGC-3') [13]. PCR reactions were performed in a final volume of 50 µL containing 5 µL 10 × buffer + MgCl2, 2 mM dNTP, 2 unit Taq DNA polymerase, 100 ng genomic DNA as a template, and 25 picomole of each primer. PCR was performed using a thermal cycler (Eppendorf Co., Germany) under the following conditions: an initial denaturation for 2 min at 94 °C; 30 cycles of 95 °C for 30 s, 60 °C for 30 s, and 72 °C for 30 s and a final extension at 72 °C for 8 min.
Genotyping of vacA gene of Helicobacter pylori
Presence of the genotypes of vacA alleles (s1a, s1b, s1c, m1a, m1b and m2) were determined by PCR. The primer sequences are shown in [14].
The PCR were performed in a total volume of 50 μL containing 1 μM of each primers, 1 μL of genomic DNA (approximately 200 ng), 1 mM of dNTPs mix (invitrogen), 2 mM of Mgcl2, and 0.05 U/μL Taq DNA polymerase (invitrogen). PCR amplifications were performed in an automated thermal cycler (Biometra Co., Germany). The following cycle conditions were used for PCR amplification: 32 cycles of 45 s at 95 °C, 50 s at 64 °C, and 70 s at 72 °C. All runs included one negative DNA control consisting of PCR grade water and two or more positive controls (26695, J99, SS1, Tx30, 88-23 and 84-183).
Gel electrophoresis
The PCR amplification products (10 μL) were subjected to electrophoresis in a 1 % agarose gel in 1X TBE buffer at 80 V for 30 min, stained with ethidium bromide, and images were obtained in a UVIdoc gel documentation systems (UK). The PCR products were identified by 100 bp DNA size marker (Fermentas, Germany).
Statistical analysis
Data were transferred to Microsoft Excel spreadsheet (Microsoft Corp., Redmond, WA, USA) for analysis. Using SPSS 16.0 statistical software (SPSS Inc., Chicago, IL, USA), Chi-square test and Fisher’s exact two-tailed test analysis was performed and differences were considered significant at p < 0.05. Distribution of H. pylori genotypes isolated from food items was statistically analyzed.
Results
Prevalence of Helicobacter pylori in various types of food items
All of the food items samples were tested using the culture and PCR methods. Total distribution of H. pylori in various types of food items samples is shown in . Of 300 food items, 60 (20 %) were positive for H. pylori. All the food items were contaminated with H. pylori. The most commonly contaminated samples were vegetable sandwich (45 %), minced meat (32 %), and meat sandwich (20 %). There were statistically significant differences amongst the incidence of bacteria in vegetable sandwich and chicken sandwich (p = 0.017), minced meat and ham (p = 0.025) and vegetable sandwich and ham (p = 0.023).
VacA genotyping pattern
Figures 1, 2 and 3 show the results of gel electrophoresis for vacA genotypes of H. pylori strains of food samples. Total distribution of vacA genotypes in the H. pylori isolates of various types of food items is shown in table 3. Results indicated that the most commonly detected genotypes were vacA s1a (66.66 %), vacA m2 (56.66 %) and vacA m1a (51.66 %). The most commonly detected genotypes in the meat based food items (minced meat, meat sandwich and ham), vegetable sandwich and ready to eat fish were vacA s1a, vacA m1a and vacA m2, respectively. Significant differences were found between the incidence of s1a and s2 (p =0.017) and also between s1a and s1c (p = 0.020), s1a and m1b (p = 0.024), m2 and s1c (p =0.018), m1a and m1b (p = 0.016), m2 and s2 (p =0.022) and m2 and s1b (p =0.025) genotypes.
Combined genotype status
Twenty-one different genotypic combinations were found for the H. pylori strains of studied food samples (). The most commonly detected combined genotypes were s1am2 (45 %), s1am1a (40 %) and m1am2 (35 %). There were no positive results for m1as2, m1bs1c, m1bs2, s1as2, s1bs2 and s1cs2 genotype.
Discussion
As far as we know, this is the first report of genotyping of H. pylori in the various types of food items in Iran and even other parts of the world. Our results showed high prevalence of H. pylori in various types of food items (20 %). The prevalence of H. pylori in our study (20 %) was higher than those of milk (12.5 %) [15], dairy products (19.2 %) [5], salad (10.86 %) [4], vegetable (13.68 %) [7]. In addition to vegetable sandwich (45 %), meat based food items including minced meat (32 %), meat sandwich (20 %) and ham (8.33 %) had high prevalence of H. pylori.
One possible explanation for the high prevalence of bacteria in vegetable sandwich is the fact that there was no sufficient time for well washing of vegetables used in the sandwich. In addition, vegetables are mainly growth in the context of animal manure and were irrigated using the contaminated water. Role of animal feces as a source for transmission of H. pylori has been reported previously [13]. High amount of activated water (AW), optimum levels of salt and pH and finally the role of infected staffs as sources of H. pylori infection [9,12] are other reasons for the high prevalence of H. pylori in vegetable sandwich samples.
Mhaskar et al [16] reported the high prevalence of peptic ulcer and H. pylori infection in patients associated with meat and meat products and restaurant foods which confirm the results of our study. High amount of Aw in meat base food samples, optimum pH and salt levels, presence of appropriate levels of amino acids including arginine, histidine, isoleucine, leucine, methionine, phenylalanine, alanine, valine, proline, serine, and tryptophan favors the for growth of H. pylori [12], this is in addition to unsanitary conditions in their preparation and cross contamination of these foods due to handling by factory and food units staffs. High prevalence of H. pylori in foods with animal origin has been reported previously [5,9,15,17,18,20].
Roles of contaminated water and even drinking water in transmission of H. pylori have been confirmed previously [17-20]. Probably, the water sources used for production, washing and processing of these foods samples were contaminated. Finally, unsanitary conditions, handling contamination, use of contaminated equipment and lack of public and individual hygiene could be the main reasons for the high prevalence of H. pylori in food samples of our study. Food safety regulations as well as quality standards – including hazard analysis and critical control points (HACCP), good agricultural practices (GAPs) and good manufacturing practices (GMPs) – should be introduced in Iranian food units to prevent proliferation of H. pylori in food samples.
Similar to the conducted studies on the clinical samples [8-10,21], the results of our investigation showed the high prevalence of vacA s1a (66.66 %), vacA m2 (56.66 %), vacA m1a (51.66 %), vacA s1am2 (45 %), vacA s1am1a (40 %) and vacA m1am2 (35 %) genotypes in food samples. High prevalence of these genotypes have been reported from the various types of foods [4,5,7] and even animals [8]. Yahaghi et al [7] reported that the vacA s1a (37.28 %) and vacA m1a (30.50 %) had the highest prevalence of genotypes in the H. pylori of vegetable and salad samples, while vacA s1c (10.16 %) had the lowest prevalence which was similar to our findings.
It seems that differences in the sources of infection in vegetable sandwich, ready to eat fish samples and meat based food items in our study, could have been the main factor for the differences in the prevalence of genotypes of H. pylori. The vacA s1a was the most commonly detected genotypes in the meat based food items which maybe showed that these samples were secondary infected (handling of samples by staffs of the factories).
In the case of vegetable sandwiches the vacA m1a was the most commonly detected which maybe showed that contact with contaminated water and animal manure are the main factors for primary infection of vegetables used in their preparation. It seems that in the ready to eat fish samples the most prevalent genotype was vacA m2 and it may be related to the type of H. pylori strains detected in this type of samples. Since H. pylori isolates in our study harbored vacA genotypes, consumption of food items contaminated with virulent strains may provoke duodenal ulceration, gastric mucosal atrophy and gastric cancer.
Conclusion
Some food items in Iran harbor H. pylori similar in the genotype of vacA alleles with isolates recovered from human clinical samples. High prevalence of H. pylori in food items suggests that contaminated foods may be the sources of the bacteria that entered the human population in Iran over a period of time. Diversity of H. pylori genotypes between various types of food samples may be indicative of the various sources of contamination of these food items.
Declarations
Acknowledgement
References
Archives


News Updates